Revista Biomķdica Revisada Por Pares

En esta conferencia se hablarß acerca de la oxidaci¾n grasa en el m·sculo esquelķtico, c¾mo se altera este proceso en la obesidad y quķ tipo de intervenciones se pueden efectuar para mejorarlo.
La captaci¾n de glucosa por el m·sculo esquelķtico se ha estudiado con tecnologĒa PET (TomografĒa de Emisi¾n de Positrones), que ha permitido demostrar que existe una flexibilidad metab¾lica en la utilizaci¾n de la glucosa, con mĒnimas tasas de captaci¾n y acumulaci¾n de glic¾geno durante el ayuno y gran incremento de estas tasas en respuesta a la insulina. En estado de ayuno, el m·sculo esquelķtico tiene una captaci¾n muy elevada de ßcidos grasos: la fracci¾n que se extrae en una ·nica pasada por el m·sculo esquelķtico es de alrededor de 40%, lo cual es muy elevado en comparaci¾n con el 1% de extracci¾n de glucosa en las mismas condiciones; esta dependencia de la grasa del m·sculo esquelķtico permite al organismo ahorrar glucosa en beneficio del cerebro y evitar la hipoglicemia durante el ayuno y el ejercicio.
La gran dependencia muscular de la oxidaci¾n grasa como fuente de energĒa en el ayuno normal se refleja en los valores del cuociente respiratorio (CR), que es el resultado de la relaci¾n entre la producci¾n de di¾xido de carbono y el consumo de oxĒgeno e indica cußl es el combustible que el organismo estß utilizando en un momento determinado. Su valor es bajo en el ayuno normal, lo que indica que se estß quemando principalmente grasa; en cambio, en los individuos con diabetes mellitus tipo 2, a pesar de que en su mayorĒa tienen sobrepeso y niveles elevados de ßcidos grasos circulantes, el cuociente se encuentra elevado, debido a que existe un defecto en la oxidaci¾n de las grasas.
En la Figura 1 se ilustra lo que ocurre en condiciones fisiol¾gicas normales, en individuos delgados, con el CR medido en la pierna (LegRQ) con la tķcnica de RQ (clamp), es decir, cambios en el CR durante un clamp. El CR es muy bajo en el ayuno, lo que representa la dependencia de la oxidaci¾n grasa, pero a medida que se infunde insulina, se estimula la captaci¾n de glucosa, se suprime la oxidaci¾n grasa y el m·sculo empieza a depender casi exclusivamente de la oxidaci¾n de la glucosa, de modo que el CR se desplaza desde un valor inicial de 0,8 a un valor aproximado de 1,0.
En la diabetes tipo 2 se observan dos anormalidades: primero, durante el estado de ayuno no existe dependencia de la oxidaci¾n grasa, fen¾meno que resulta fascinante, precisamente porque hay gran cantidad de grasa disponible para la combusti¾n, lo que implica que debe haber alg·n defecto dentro del tejido, ya sea para obtener la grasa desde el plasma, que en realidad no es el caso, o para efectuar la distribuci¾n intracelular de la grasa para la oxidaci¾n. En segundo lugar, este defecto en la utilizaci¾n de la grasa persiste frente a la estimulaci¾n con insulina. En la Figura 2 se ilustra la pķrdida de la adaptaci¾n, de la homeostasis y de la capacidad de respuesta a las condiciones de ayuno y de estimulaci¾n con insulina que se observa en la diabetes.
Figura 2. CalorimetrĒa regional indirecta (Kelley, Mandarino. J Clin Invest 86, 1999-2007, 1990).
En los obesos existe una inflexibilidad metab¾lica, que se ilustra en la Figura 3, donde se demuestra que el m·sculo normal tiene una gran capacidad para hacer una transici¾n de su patr¾n de oxidaci¾n de combustible, dependiendo de si el sujeto acaba de comer o si estß en ayunas, mientras que en la obesidad se pierde esta capacidad homeostßtica. El mķdico internista siempre piensa en las enfermedades cr¾nicas como entidades en las que ocurre una pķrdida de las reservas fisiol¾gicas: tanto la insuficiencia cardiaca congestiva como el enfisema o la insuficiencia renal cr¾nica se caracterizan por una pķrdida crĒtica de la capacidad homeostßtica; en este contexto, tambiķn se debe pensar en la resistencia insulĒnica que aparece en la obesidad, como en una pķrdida de la reserva fisiol¾gica, en cuanto a la capacidad para cambiar entre los diferentes combustibles.
Para profundizar en el significado de estas observaciones fisiol¾gicas se decidi¾ efectuar el anßlisis bioquĒmico de las vĒas del catabolismo graso, en peque±os trozos de m·sculo obtenido mediante biopsia con aguja a nivel del muslo. Se midi¾ el contenido de triglicķridos en el m·sculo y la actividad de los distintos sistemas enzimßticos que participan en la descomposici¾n y oxidaci¾n grasa, que en su mayorĒa corresponden al complejo enzimßtico carnitin-palmitil transferasa (CPT), responsable de remover los ßcidos grasos de cadena larga desde el citosol a la matriz mitocondrial para la oxidaci¾n. Los resultados indicaron que la actividad de la CPT estß disminuida en el m·sculo de los individuos obesos en comparaci¾n con los delgados; tambiķn estßn disminuidas las enzimas que participan en la vĒa de la oxidaci¾n beta, en la que se descomponen los ßcidos grasos a acetil-CoA para que puedan entrar al ciclo de Krebs, de modo que la actividad de la citrato sintetasa, cuya actividad es un marcador del ciclo de Krebs, estß reducida, al igual que la actividad de la beta-hidroxi-acil-dehidrogenasa, otra enzima marcadora de la cadena transportadora de electrones. En conjunto, esto se traduce en una disminuci¾n de la capacidad enzimßtica del m·sculo esquelķtico para efectuar el catabolismo de los ßcidos grasos, lo que predispone al m·sculo a almacenar grasa en lugar de oxidarla. AsĒ se explica que el contenido graso del m·sculo se encuentre aumentado en la obesidad, como se ha demostrado mediante varias tķcnicas.
En la Figura 4 se muestra el resultado de estudios histol¾gicos efectuados mediante la tinci¾n con aceite rojo O (ORO), que ti±e la grasa y permite verla como gotas rojas dentro de las fibras musculares, que tambiķn se pueden observar con tomografĒa computada, en forma no invasiva, o con espectroscopĒa de resonancia magnķtica, entre muchas otras tķcnicas que se pueden utilizar. Todas ellas muestran que el contenido de grasa dentro del m·sculo estß aumentado en la resistencia a la insulina, la obesidad y la diabetes. Se puede observar que el contenido graso de las fibras musculares es varias veces mßs elevado en la obesidad y en la diabetes tipo II; las barras blancas demuestran que la baja de peso moderada, entre 10 y 15%, se asocia a reducci¾n de la grasa intramuscular.
En cuanto a la causa de estas anomalĒas en la oxidaci¾n grasa, el m·sculo esquelķtico humano estß compuesto por varios tipos de fibras, siendo las principales las fibras tipo I, oxidativas, de contracci¾n lenta, de las que depende la oxidaci¾n grasa, y las fibras tipo II, mßs bien glicolĒticas, de contracci¾n rßpida. Durante mucho tiempo se plante¾ que las personas que estßn predispuestas a desarrollar obesidad deberĒan tener reducido el n·mero de fibras tipo I y aumentado el n·mero de fibras tipo II, lo que traerĒa como consecuencia que ese m·sculo serĒa menos capaz de quemar grasa. Para confirmar esta hip¾tesis se estudiaron muestras humanas de fibras musculares, con el objetivo de determinar el contenido graso de cada tipo de fibra. En la Figura 5 se muestran secciones seriadas de una biopsia muscular, en las que se han hecho tinciones sucesivas para determinar, primero, el tipo de fibra (ATP); en la siguiente tinci¾n, para analizar el contenido graso (Oil Red O); y en las siguientes, para ver la actividad de la succinato deshidrogenada (SDH) y la GPDH, enzimas que participan en la cadena transportadora de electrones y que son un Ēndice de la actividad mitocondrial. Se observan cuatro secciones de tres tipos de individuos: delgado, obeso y diabķtico tipo II, en las que se puede ver que el contenido graso tiende a aumentar y la actividad mitocondrial disminuye.
Figura 5. Relaci¾n entre tipo de fibra y contenido graso.
El anßlisis computacional de las imßgenes permite cuantificar el contenido graso; se esperarĒa que las fibras tipo I tengan mßs grasa que las fibras tipo II y precisamente eso es lo que se observa, pero dentro de cada tipo de fibra, los individuos que son obesos y tienen diabetes tipo II tienen mßs grasa en las fibras tipo I, tipo II y tipo IIb. En relaci¾n a la actividad mitocondrial, por definici¾n se esperarĒa que las fibras tipo I tuvieran alto grado de actividad, en comparaci¾n con las tipo II, y eso es exactamente lo que se observa, pero los individuos con obesidad y diabetes tipo II tienen disminuci¾n de la actividad mitocondrial dentro de cada tipo de fibra (Figura 6).
Esto llev¾ a analizar con mßs detalle la actividad mitocondrial, para ver si existe diferencia entre la obesidad y la diabetes tipo 2. En la Figura 7 se muestra una vista de microscopĒa electr¾nica de una mitocondria entre las miofibrillas del m·sculo esquelķtico, junto a una peque±a gota de grasa, que es un combustible importante; esta morfologĒa es bastante distinta en la fibra muscular del diabķtico tipo 2, a la derecha, en la cual las mitocondrias son mßs peque±as y la grasa es mßs abundante.
Figura 7. Disfunci¾n mitocondrial en la resistencia insulĒnica (Kelley et al, Diabetes 2002).
Se ha demostrado que el menor tama±o mitocondrial se correlaciona con la gravedad de la resistencia a insulina. En el grßfico de la Figura 8 se muestra la habilidad de la insulina para estimular el metabolismo graso dentro del organismo; como se puede ver, los individuos que tienen una elevada sensibilidad a la insulina tienden a tener mitocondrias de mayor tama±o que aquellos que presentan resistencia a la insulina.
De la misma manera, al determinar la actividad de la NADH-oxidasa en mitocondrias aisladas del m·sculo esquelķtico, se observa que el menor tama±o mitocondrial se asocia a menor actividad de esta enzima. Se sabe que ķsta es un reflejo de la actividad de la cadena transportadora de electrones, la que disminuye significativamente en la obesidad, mßs a·n si se agrega la diabetes tipo 2 (vķase Figura 9). Este grado de disminuci¾n no es lo que se ve habitualmente en las enfermedades hereditarias de la mitocondria, en que los defectos son mucho mßs marcados, es decir, estos no son defectos hereditarios mayores autos¾micos recesivos o dominantes, propios de la enfermedad mitocondrial; pero estas disfunciones de 20% a 30% exponen al m·sculo a la acumulaci¾n de grasa con el transcurso del tiempo.
En los ·ltimos a±os hemos intentado estudiar la resistencia a la insulina desde la perspectiva de la biologĒa celular, para comprender la naturaleza del da±o a nivel mitocondrial. Una posibilidad es que exista disminuci¾n del n·mero de mitocondrias, lo que se puede determinar midiendo el contenido de ADN mitocondrial, en relaci¾n al contenido de ADN nuclear dentro del m·sculo, Ēndice que parece estar reducido en forma significativa, alrededor de 30%. Sin embargo, al analizar la actividad funcional de la mitocondria se observa una reducci¾n mayor que lo que corresponde a la reducci¾n del contenido de ADN mitocondrial; por lo tanto, ademßs de una reducci¾n en el n·mero de mitocondrias habrĒa una disfunci¾n mitocondrial (Ritov et al, Diabetes 2005).
Tambiķn se puede plantear la existencia de dos poblaciones diferentes de mitocondrias. En la Figura 10 se muestran las que se ubican justo por debajo de la membrana celular, llamadas mitocondrias del subsarcolema. Muchas de ellas se agrupan alrededor del n·cleo de la cķlula muscular, proveyendo energĒa para ķste; otras se disponen a lo largo de la superficie celular y estßn involucradas en el intercambio i¾nico y tambiķn en el transporte de sustratos, asĒ como en la activaci¾n y transporte de glucosa. Un segundo grupo de mitocondrias son las mitocondrias intramiofibrilares, que se ubican alrededor de las fibras musculares y proveen la energĒa para la contracci¾n muscular; ķstas se pueden estudiar por separado usando tķcnicas como la microscopĒa electr¾nica o tķcnicas bioquĒmicas de aislamiento. En la Figura 10 se observa una capa gruesa de mitocondrias ubicadas en la zona subsarcolķmica en un individuo delgado, mientras que en el diabķtico tipo 2, al igual que en el obeso, se observa una reducci¾n muy marcada de las mitocondrias a lo largo de la superficie celular. De la misma manera, existe un dķficit de mitocondrias dentro de la poblaci¾n intramiofibrilar. Mßs tarde se se±alarß la importancia de este hallazgo en la patogenia de la resistencia a la insulina.
Figura 10. Las mitocondrias subsarcolķmicas estßn disminuidas en la diabetes tipo 2 y la obesidad.
Otro enfoque consiste en hacer anßlisis de ordenamiento genķtico (DNA microarrays). En los ·ltimos a±os han aparecido varias publicaciones importantes sobre la relaci¾n entre los patrones de expresi¾n genķtica en el m·sculo esquelķtico en individuos con diabetes tipo 2 y en individuos que estßn en riesgo de presentar esta enfermedad. Uno de los hallazgos interesantes que ha emergido de esos anßlisis es que el patr¾n mßs consistente es compatible con una alteraci¾n de la expresi¾n de los genes de la fosforilaci¾n oxidativa, lo que reafirma el concepto de que la resistencia a la insulina tiene que ver con un desequilibrio energķtico (Vamsi K Mootha et al. Nature Genetics, June 2003). En esta instancia, se producen dentro del m·sculo algunas anomalĒas sutiles, que despuķs de un largo tiempo de exposici¾n del tejido a la acumulaci¾n de la grasa, generan resistencia a la insulina.
Por supuesto que la labor del mķdico consiste no s¾lo en hacer un buen diagn¾stico y comprender la fisiopatologĒa, sino que ademßs debe intervenir e idear tratamientos eficaces. En este contexto, nuestro grupo ha efectuado algunos estudios adicionales para determinar los efectos de la baja de peso por sĒ sola, sin actividad fĒsica, y de la pķrdida de peso combinada con actividad fĒsica, usando un programa similar al que se ha utilizado en el Programa de Prevenci¾n en Diabetes realizado en los Estados Unidos y que consiste en realizar ejercicio de moderada intensidad y lograr una baja de peso de 5 a 10% del peso inicial. Ademßs se analiz¾ un tercer grupo en que se intervino solamente con ejercicio.
Este ·ltimo estudio fue realizado por Goodpaster en ancianos y demostr¾ muy buenos efectos sobre el m·sculo esquelķtico, ya que el contenido de ADN mitocondrial aument¾ en 60%, al igual que la actividad de la cadena transportadora de electrones y la cardiolipina. Es necesario recordar que la cardiolipina es un fosfolĒpido ·nico de la membrana interna mitocondrial, de tal modo que su contenido refleja la cantidad de membrana mitocondrial interna, que es la que contiene la cadena transportadora de electrones y los componentes que producen energĒa; es una buena forma de cuantificar la cantidad de mitocondrias en una muestra de biopsia muscular. En este estudio, los individuos aumentaron la oxidaci¾n grasa durante el ejercicio y presentaron proliferaci¾n mitocondrial, respuesta que probablemente es importante para bajar de peso y mßs a·n, para mantener la pķrdida de peso.
Frente a la baja de peso sola, sin actividad fĒsica asociada, ocurren varias cosas interesantes desde el punto de vista del metabolismo corporal total, pero en tķrminos de la capacidad de oxidaci¾n grasa el impacto no es tan bueno. Disminuye el contenido de lĒpidos intramiocelular, como parte de la baja total de peso, porque mejora el metabolismo de la glucosa y la respuesta a la insulina; pero no hay un cambio en la cantidad de mitocondrias ni en la capacidad de oxidaci¾n grasa (Figura 11). Esto quiere decir que, a menos que estos individuos puedan mantener una ingesta cal¾rica restringida, van a estar muy propensos a volver a almacenar grasa dentro del m·sculo; es decir, tienen un riesgo importante de volver a ganar el peso perdido. Como ya se ha se±alado, mejoran la oxidaci¾n de la glucosa en respuesta a la insulina, o sea, mejoran la tolerancia a la glucosa y la hiperglicemia.
Figura 11. Efecto de la pķrdida de peso, sin actividad fĒsica.
En otro grupo se hizo el mismo tipo de intervenci¾n, pero esta vez se solicit¾ a los individuos que caminaran entre media a una hora a velocidad moderada, ademßs de intentar bajar de peso. Esta intervenci¾n fue exitosa, ya que los sujetos aumentaron su capacidad aer¾bica en 15%, bajaron su peso en 7% respecto al peso inicial y aumentaron su sensibilidad a la insulina en 40%, cifra realmente notable, ya que no existe ning·n medicamento que tenga un efecto tan poderoso sobre la resistencia a la insulina. Lo importante es que se observ¾ aumento en el CR en respuesta a la insulina y disminuci¾n de este parßmetro en respuesta al ayuno, lo que implica que mejor¾ la capacidad para quemar grasa en la condici¾n de ayuno. Esto se observa en la Figura 12, en la que se grafican las variables sensibilidad a la insulina y flexibilidad metab¾lica en el rango de CR que va desde el ayuno a la estimulaci¾n con insulina, y se observa que estas dos variables estßn fuertemente relacionadas. A medida que se recupera la flexibilidad metab¾lica, esto va de la mano con la recuperaci¾n de la sensibilidad a la insulina.
Posteriormente se trat¾ de determinar cußles eran los mejores predictores de mejorĒa de la sensibilidad a la insulina: la adiposidad visceral, la disminuci¾n generalizada de la masa grasa o el cambio en el consumo mßximo de oxĒgeno (capacidad aer¾bica); y se encontr¾ que todos esos parßmetros se correlacionaban con cambios en la sensibilidad a la insulina, pero el factor mßs fuerte fue el aumento de la oxidaci¾n grasa en condiciones basales, lo que refuerza el hecho de que la actividad fĒsica es extremadamente importante en el tratamiento de la obesidad, ya que va a producir un aumento en la capacidad de oxidaci¾n grasa. En la Figura 13 se observa c¾mo las mitocondrias han aumentado de tama±o despuķs de la actividad fĒsica de moderada intensidad, lo que se correlacion¾ con una mejorĒa en la sensibilidad a la insulina.
Figura 13. Mitocondrias del m·sculo esquelķtico antes y despuķs de la intervenci¾n.
En cuanto a la capacidad funcional de la mitocondria, se esperaba que aumentara el ADN mitocondrial en respuesta a la intervenci¾n, pero esto no ocurri¾; es probable que esto se deba a que el control de las mitocondrias no es ejercido en forma exclusiva por el ADN mitocondrial, ya que el ARN mensajero que se deriva del ADN mitocondrial generalmente existe en una cantidad diez veces mayor que el ARN mensajero que se deriva de los genes del n·cleo que codifican para la oxidaci¾n. En otras palabras, las mitocondrias viven dentro de la cķlula muscular, colonizan esa cķlula, pero el control sobre ellas es ejercido por el n·cleo. En cambio, la actividad de la citrato sintetasa, la enzima de la matriz que participa en el ciclo de Krebs, aument¾ significativamente y hubo un gran incremento del contenido de cardiolipina y de la actividad de NADH oxidasa y SDh, enzimas de la cadena transportadora de electrones. Se puede decir que la mitocondria present¾ todas estas adaptaciones para mejorar la oxidaci¾n de las grasas.
En cuanto a las mediciones de actividad lipĒdica, ya se se±al¾ c¾mo la caĒda del CR en el ayuno se correlaciona con un aumento de la sensibilidad a la insulina. El contenido graso del m·sculo, que te¾ricamente debĒa disminuir, se mantuvo mßs o menos igual. Este aspecto es muy interesante: si se observa el m·sculo de atletas de alta resistencia, como los corredores de cross-country o los monta±istas, se encuentra que tienen un alto contenido de grasa en su m·sculo, pero tambiķn tienen mitocondrias muy grandes y activas, de modo que esa grasa es utilizada como un combustible de rßpido recambio, que se deposita por el aporte de la dieta, pero se quema rßpidamente, a diferencia de lo que ocurre en los individuos obesos, en los que el dep¾sito de grasa es estßtico, porque la capacidad para quemarlo es mucho menor. A medida que aumenta la actividad la cantidad de grasa en el m·sculo permanece mßs o menos igual, pero el tama±o de las gotas de grasa se reduce, como se demostr¾ con tķcnicas especiales de microscopĒa; es decir, tienen la misma cantidad de grasa, pero esparcida en muchas gotas mßs peque±as, que se acercan a la mitocondria y se recambian mßs rapidamente. El cambio en el tama±o de las gotas de lĒpidos se correlacion¾ con una mejorĒa en la sensibilidad a la insulina.
Un ßrea de investigaci¾n muy interesante es el estudio de la biologĒa de estas gotas de lĒpidos. En un ejemplar de Science del a±o 2006 se se±ala que las gotas lipĒdicas, que antes se consideraban como meras acumulaciones de grasa en ciertas zonas, son probablemente los organelos endocrinos mßs activos del organismo, ya que parecen estar involucrados en la sĒntesis del colesterol y en muchos otros procesos. Es posible que estas peque±as gotas de grasa dentro de las cķlulas contengan gran cantidad de informaci¾n y sitios especĒficos de regulaci¾n; y que cuando se aclare esto se encuentren nuevos blancos de intervenci¾n. En este contexto, puede ser importante saber que no s¾lo la cantidad de grasa dentro del m·sculo se altera por el tipo de enfermedad, sino tambiķn el tama±o de las gotas de grasa, que es un poco mayor en la diabetes y en la obesidad, en comparaci¾n con los individuos delgados.
Para finalizar, es muy interesante analizar la distribuci¾n de las mitrocondrias en el m·sculo y su relaci¾n con la resistencia a la insulina, en tķrminos de aumentar el transporte de glucosa en respuesta a la se±al que da esta hormona, lo que define la capacidad del m·sculo para absorber glucosa y determina la sensibilidad a la insulina. Pues bien, la forma en que la mitocondria podrĒa estar involucrada en la resistencia a la insulina es a travķs de la acumulaci¾n de grasa dentro del m·sculo, acumulaci¾n que interferirĒa con la se±al de la insulina.
Por otra parte, a medida que la glucosa entra a la cķlula como glucosa libre, debe ser capturada para ser transformada en glucosa 6 fosfato; de lo contrario quedarĒa libre para abandonar la cķlula por la misma puerta por la que entr¾. La hexoquinasa, que estß unida a la mitocondria justo en el canal de porina, es muy importante en la bioenergķtica de la mitocondria, porque mantiene ese estado libre de fosforilaci¾n oxidativa, eficiente entre mitocondrias pareadas. Esta hexoquinasa estß tĒpicamente disminuida en el m·sculo esquelķtico de personas con diabetes; una vez mßs, si hay escasez de mitocondrias cercanas a la superficie celular donde estß ocurriendo el transporte de glucosa, esto tambiķn puede impedir la captaci¾n de glucosa. En esta hip¾tesis se estß trabajando en nuestro laboratorio en el ·ltimo a±o.

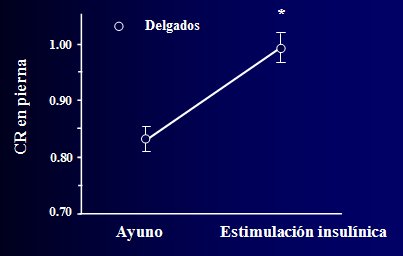 Figura 1. Flexibilidad metab¾lica de la oxidaci¾n de glucosa y lĒpidos en el individuo normal (Kelley, Goodpaster, Wing, Simoneau, AJP 1999).
Figura 1. Flexibilidad metab¾lica de la oxidaci¾n de glucosa y lĒpidos en el individuo normal (Kelley, Goodpaster, Wing, Simoneau, AJP 1999).
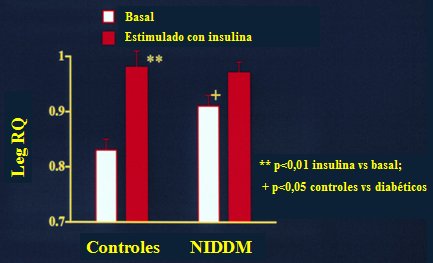 Figura 2. CalorimetrĒa regional indirecta (Kelley, Mandarino. J Clin Invest 86, 1999-2007, 1990).
Figura 2. CalorimetrĒa regional indirecta (Kelley, Mandarino. J Clin Invest 86, 1999-2007, 1990).
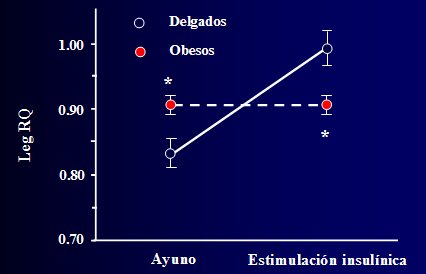 Figura 3. Inflexibilidad metab¾lica de la oxidaci¾n de glucosa y lĒpidos en la obesidad (Kelley, Goodpaster, Wing, Simoneau, AJP 1999).
Figura 3. Inflexibilidad metab¾lica de la oxidaci¾n de glucosa y lĒpidos en la obesidad (Kelley, Goodpaster, Wing, Simoneau, AJP 1999).
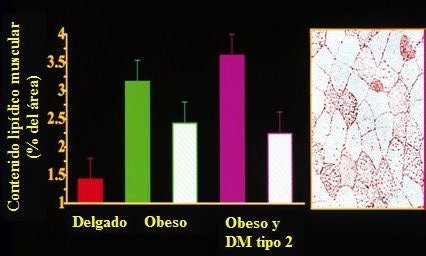 Figura 4. El contenido de grasa de la fibra del m·sculo esquelķtico estß aumentado en la obesidad y disminuye con la pķrdida de peso.
Figura 4. El contenido de grasa de la fibra del m·sculo esquelķtico estß aumentado en la obesidad y disminuye con la pķrdida de peso.
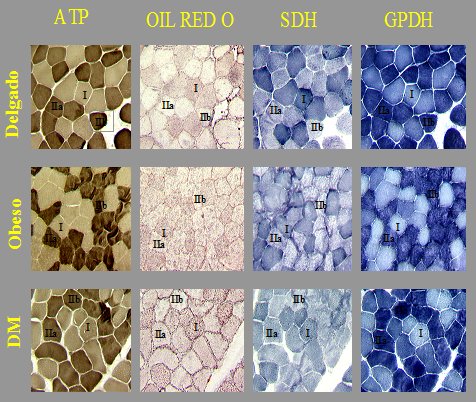 Figura 5. Relaci¾n entre tipo de fibra y contenido graso.
Figura 5. Relaci¾n entre tipo de fibra y contenido graso.
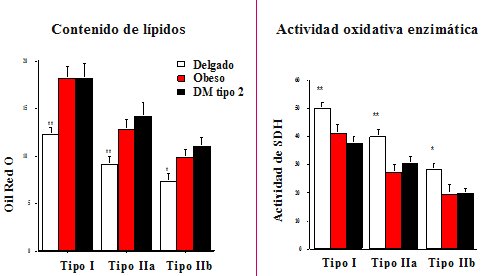 Figura 6. Los lĒpidos intra-miocelulares estßn aumentados en todos los tipos de fibra en la obesidad y la DM tipo 2 y la actividad enzimßtica oxidativa estß disminuida.
Figura 6. Los lĒpidos intra-miocelulares estßn aumentados en todos los tipos de fibra en la obesidad y la DM tipo 2 y la actividad enzimßtica oxidativa estß disminuida.
 Figura 7. Disfunci¾n mitocondrial en la resistencia insulĒnica (Kelley et al, Diabetes 2002).
Figura 7. Disfunci¾n mitocondrial en la resistencia insulĒnica (Kelley et al, Diabetes 2002).
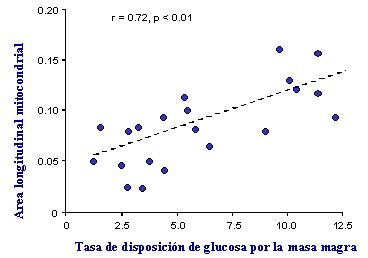 Figura 8. El tama±o de las mitocondrias, medido por microscopĒa electr¾nica, se relaciona con la resistencia a la insulina (Kelley et al, Diabetes 2002).
Figura 8. El tama±o de las mitocondrias, medido por microscopĒa electr¾nica, se relaciona con la resistencia a la insulina (Kelley et al, Diabetes 2002).
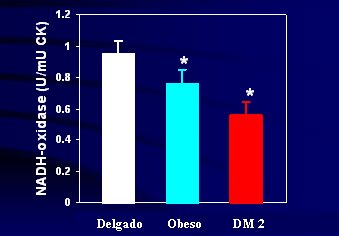 Figura 9. Actividad de NADH-oxidasa en el m·sculo esquelķtico en individuos delgados no diabķticos, obesos no diabķticos y diabķticos tipo 2 (Kelley et al, Diabetes 2002).
Figura 9. Actividad de NADH-oxidasa en el m·sculo esquelķtico en individuos delgados no diabķticos, obesos no diabķticos y diabķticos tipo 2 (Kelley et al, Diabetes 2002).
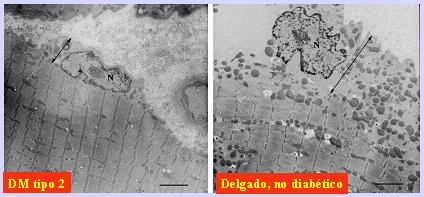 Figura 10. Las mitocondrias subsarcolķmicas estßn disminuidas en la diabetes tipo 2 y la obesidad.
Figura 10. Las mitocondrias subsarcolķmicas estßn disminuidas en la diabetes tipo 2 y la obesidad.
 Figura 11. Efecto de la pķrdida de peso, sin actividad fĒsica.
Figura 11. Efecto de la pķrdida de peso, sin actividad fĒsica.
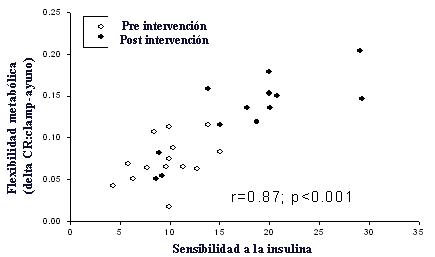 Figura 12. Flexibilidad metab¾lica y sensibilidad a la insulina antes y despuķs de la intervenci¾n (ejercicio + baja de peso).
Figura 12. Flexibilidad metab¾lica y sensibilidad a la insulina antes y despuķs de la intervenci¾n (ejercicio + baja de peso).
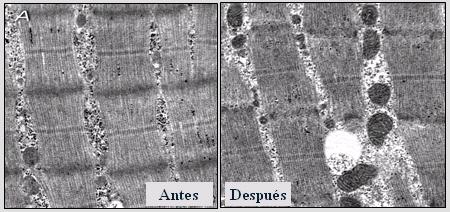 Figura 13. Mitocondrias del m·sculo esquelķtico antes y despuķs de la intervenci¾n.
Figura 13. Mitocondrias del m·sculo esquelķtico antes y despuķs de la intervenci¾n.
 Esta obra de Medwave estß bajo una licencia Creative Commons Atribuci¾n-NoComercial 3.0 Unported. Esta licencia permite el uso, distribuci¾n y reproducci¾n del artĒculo en cualquier medio, siempre y cuando se otorgue el crķdito correspondiente al autor del artĒculo y al medio en que se publica, en este caso, Medwave.
Esta obra de Medwave estß bajo una licencia Creative Commons Atribuci¾n-NoComercial 3.0 Unported. Esta licencia permite el uso, distribuci¾n y reproducci¾n del artĒculo en cualquier medio, siempre y cuando se otorgue el crķdito correspondiente al autor del artĒculo y al medio en que se publica, en este caso, Medwave.
Expositor:
David Kelley[1]
Citaci¾n: Kelley D. Metabolic disorders of fatty acids in muscle tissue and fat in obese patients. Medwave 2007 Ene;7(1):e2370 doi: 10.5867/medwave.2007.01.2370
Fecha de publicaci¾n: 1/1/2007
Nos complace que usted tenga interķs en comentar uno de nuestros artĒculos. Su comentario serß publicado inmediatamente. No obstante, Medwave se reserva el derecho a eliminarlo posteriormente si la direcci¾n editorial considera que su comentario es: ofensivo en alg·n sentido, irrelevante, trivial, contiene errores de lenguaje, contiene arengas polĒticas, obedece a fines comerciales, contiene datos de alguna persona en particular, o sugiere cambios en el manejo de pacientes que no hayan sido publicados previamente en alguna revista con revisi¾n por pares.
A·n no hay comentarios en este artĒculo.
Para comentar debe iniciar sesi¾n







 Estudios originales
Estudios originales